Immunantworten gegen Krankheitserreger werden in das angeborene und das adaptive Immunsystem unterteilt. Die adaptive Immunantwort, auch erworbenes Immunsystem genannt, besteht aus 2 Hauptmechanismen: der humoralen und zellvermittelten Immunantwort. Die humorale Immunantwort wird durch B-Zellen (die Antikörper produzieren) vermittelt, während die zellvermittelte Immunantwort T-Zellen umfasst. Wenn das angeborene Immunsystem eine Infektion nicht ausreichend bekämpfen kann, wird die adaptive Reaktion aktiviert. Das adaptive Immunsystem ist langsamer und reagiert über einen längeren Zeitraum, aber die Aktivierung führt im Allgemeinen zur Ausbildung eines hochspezifischen immunologischen Gedächtnisses. Die 2 wichtigen Merkmale der adaptiven Reaktion sind Spezifität (mit Antigenerkennung) und Gedächtnis (Immunantwort, die für Reinfektion gespeichert wird).
Kostenloser
Download
Lernleitfaden
Medizin ➜
Das Immunsystem ist für die Abwehr (Immunität) gegen eindringende Krankheitserreger, die von Viren bis hin zu Parasiten reichen, zuständig. Seine Bestandteile sind durch Blut- und Lymphkreislauf miteinander verbunden.
Es gibt zwei sich überschneidende Komponenten:
| Angeborenes Immunsystem angeborenes Immunsystem Zellen des angeborenen Immunsystems | Adaptives Immunsystem | |
|---|---|---|
| Genetik Genetik Grundbegriffe der Genetik | Kodierung in der Keimbahn | Genveränderungen, die die Lymphozytenentwicklung beeinflussen |
| Immunantwort | Unspezifisch | Hochspezifisch |
| Zeitpunkt der Reaktion | Sofort (Minuten bis Stunden) | Entwickelt sich im Laufe der Zeit |
| Gedächtnisreaktion | Keine | Reagiert bei Antigenerkennung schnell mit Gedächtnis-Antwort |
| Erregererkennung | PRRs wie Toll-like Rezeptoren Rezeptoren Rezeptoren (Englisches Akronym: TLRs) erkennen PAMPs | |
| Komponenten |
|
|
Bei der adaptiven Immunantwort werden humorale und zellvermittelte Systeme der Immunreaktion unterschieden.

Zellvermittelte Immunität:
Die Aktivierung von T-Helferzellen führt zur Freisetzung von Zytokinen. Das Ergebnis ist die Aktivierung von zytotoxischen T-Zellen und Phagozyten (z. B. Makrophagen).

Humorale Immunantwort:
Diese Art der adaptiven Immunität wird durch B-Zellen und Antikörper vermittelt.

Die Funktionen von Antikörpern:
Antikörper haben mehrere Funktionen in unserer Immunität. Dazu gehören die Neutralisierung (von Mikroben und Toxinen), die Förderung der Phagozytose und die Aktivierung von NK-Zellen. Darüber hinaus spielen sie eine Rolle bei der Komplementaktivierung, die zur direkten Lyse von Mikroben, zur Opsonisierung und Phagozytose sowie zur Rekrutierung/Aktivierung von Neutrophilen führen kann.

Wege der Antigenpräsentation durch MHC-I- und -II-Moleküle:
Bei der Ag-Präsentation der Klasse I (links) bauen Proteasomen endogene Ags oder Proteine (innerhalb der Zelle) zu Peptiden ab. Peptidfragmente werden (über den transporter associated with antigen processing [engl. Akronym: TAP]) in das endoplasmatische Retikulum (Abkürzung: ER) transportiert, wo sie durch Aminopeptidasen weiter gekürzt und auf das MHC-I-Molekül geladen werden. Die beladenen MHC-I-Komplexe kommen zur posttranslationalen Modifikation an den Golgi-Apparat. Anschließend werden die Komplexe an die Zelloberfläche transportiert, wo sie CD8+ T-Zellen präsentiert werden. Bei der Ag-Präsentation der Klasse II (rechts) werden extrazelluläre/exogene Ags über Phagosomen von antigenpräsentierenden Zellen aufgenommen. Die Phagosomen verschmelzen dann mit dem mit proteolytischen Enzymen gefüllten Lysosomen. Dies führt zum Abbau von phagozytierten Proteinen in kleine Peptide. Währenddessen werden im ER MHC-II-Moleküle synthetisiert. Diese Moleküle haben eine invariante Kette (rosa Struktur im rechten Bild, mit „Ii“ gekennzeichnet), die an der Ag-Bindungsstelle bindet. Wenn die Bindungsstelle (durch die invariante Kette) verschlossen ist, können ER-residente Peptide nicht binden. Nach Austritt aus dem ER leitet die invariante Kette den MHC II-Komplex zum angesäuerten Endosom (wo sich die Ag-Peptide befinden). Wenn MHC-II-Komplexe an das Endosom gelangen, wird die invariante Kette freigesetzt, was das Beladen der MHC-II-Moleküle mit Ag-Peptiden (begleitet vom Protein, HLA-DM) ermöglicht. Nach der Beladung werden die gebildeten Ag-Peptid-MHC-II-Komplexe an die Zelloberfläche gebracht, um das Ag den CD4+-T-Zellen zu präsentieren.
Ii: MHC-II – assoziierte invariante Kette
MIIC: MHC-II-Kompartiment

Aktivierte T-Zellen:
Nach der Interaktion mit Ag-präsentierenden Zellen kommt es zur Stimulation und klonalen Expansion von CD4+ T-Zellen. Diese Zellen setzen Zytokine frei, die verschiedene Wirkungen haben können, wie die Aktivierung von Makrophagen (links), B-Zellen (Mitte) und CD8+ T-Zellen.
| CD4+ T-Zellen T-Zellen T-Zellen | Stimuliert durch | Produzierte Zytokine | Funktionen | Rolle bei Erkrankungen |
|---|---|---|---|---|
| Th1 | IL-12, IFN-γ | IFN-γ, TNF TNF Tumornekrosefaktor (TNF), IL-2 |
|
|
| Th2 | IL-2, IL-4 | IL-4, IL-5, IL-6, IL-9, IL-10, IL-13 |
|
|
| Th17 | IL-1, IL-6, IL-23, TGF-β | IL-17, IL-21, IL-22 | Förderung einer neutrophilen Entzündung Entzündung Entzündung |
|
| Tfh | IL-6 | IL-4, IL-21 | Erleichtern die Aktivierung und Reifung von B-Zellen B-Zellen B-Zellen | Antikörperproduktion |
| Treg | TGF-β, IL-2 | TGF-β, IL-10, IL-35 |
|
Autoimmunität Autoimmunität Autoimmunität, Allergie, Entzündung Entzündung Entzündung ↓ |
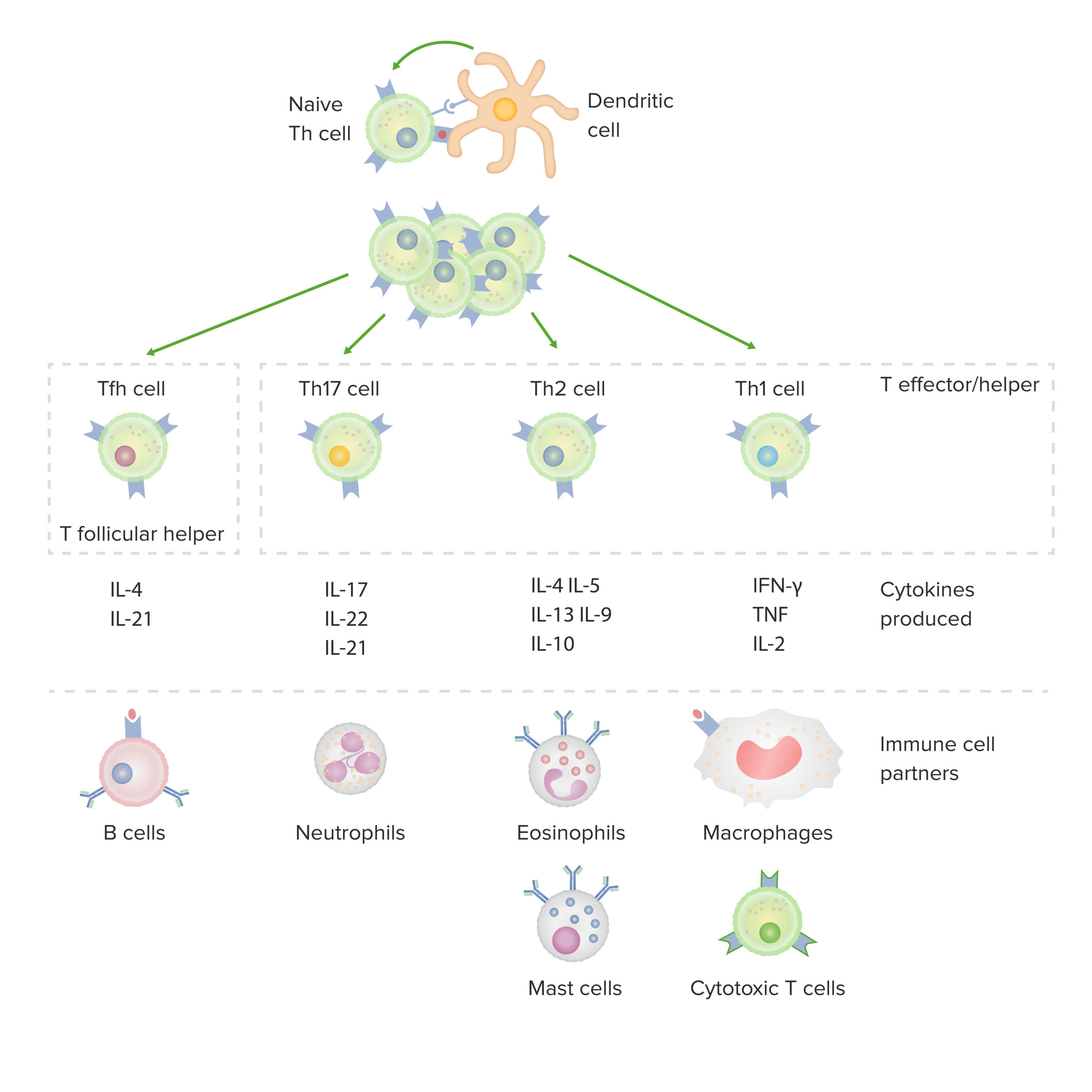
Subtypen von CD4+ T-Zellen, einschließlich der produzierten Zytokine und der damit verbundener Interaktion mit anderen Immunzellen:
IFN: Interferon
TNF: Tumor-Nekrose-Faktor

B-Zell-Rezeptor (BCR):
Besteht aus dem Immunglobulin-Molekül und dem Signalmolekül.
Das membrangebundene Ig ist an der Zelloberfläche verankert. Die Ig enthält zwei identische schwere Ketten und zwei identische leichte Ketten, die durch eine Disulfidbrücke verbunden sind.

B-Zell-Aktivierung bei der humoralen Immunantwort mit dem Ziel der Antikörperproduktion:
1) Eine B-Zelle wird durch Kontakt mit einem Ag stimuliert.
2) Sie internalisiert den Ag und zerlegt ihn.
3) Dann präsentiert sie Fragmente des Ag auf spezifischen MHC-Molekülen.
4) Der Ag-MHC-Komplex zieht T-Zellen an, die für den Reifungsprozess passend sind.
5) Von der T-Zelle sezernierte Zytokine regen die Teilung der B-Zelle und ihre Ausreifung zu einer antikörperproduzierenden Plasmazelle an.
6) Sich im Körperkreislauf frei bewegend, suchen Ak nach passenden Ags. Die Ag-Ak-Komplexe werden dann über das Komplementsystem oder von der Leber oder Milz abgebaut.
Eine Gedächtniszelle ist ein Ag-spezifischer B- oder T-Lymphozyt, der nach erneuter Exposition gegenüber demselben Erreger eine starke Immunantwort hervorruft. Sowohl B-Gedächtnis- als auch T-Gedächtniszellen exprimieren den Oberflächenmarker CD27.

Primäre und sekundäre Immunreaktion:
Bei einer primären Immunantwort werden naive B-Zellen durch ein Antigen (Ag) stimuliert. Es kommt zur B-Zell-Aktivierung und Differenzierung in antikörpersezernierende Zellen. Die Aks sind spezifisch für das auslösende Ag. Es wird erst IgM und schließlich IgG prodziert. Während der ersten Immunantwort ist die Produktion relativ gering. Bei der sekundären Immunantwort stimuliert das gleiche Ag die Gedächtnis-B-Zellen, was zur Produktion größerer Mengen spezifischer Aks führt. Produktion und Freisetzung von IgG erfolgen früher.



















