Bindegewebe ist ein Grundgewebe, das aus dem embryonalen Mesenchym stammt und mit Ausnahme von Gehirn und Rückenmark Rückenmark Rückenmark im gesamten Körper vorhanden ist. Hauptfunktion des Bindegewebes ist die strukturelle Unterstützung und mechanische Stabilität der Organe. Bindegewebe setzt sich aus einer zellulären Komponente und einer extrazellulären Matrix zusammen. Die Menge, das Verhältnis und die Zusammensetzung dieser Komponenten sind entscheidende Determinanten für die Art des Bindegewebes. Es gibt verschiedene Arten von Bindegewebe wie beispielsweise Blut, Knochen Knochen Aufbau der Knochen und Knorpel, die spezialisierte Bindegewebsformen bilden.
Kostenloser
Download
Lernleitfaden
Medizin ➜
Bindegewebe umfasst eine Gruppe von Geweben mesenchymalen Ursprungs (nicht nur ein einzelner Gewebetyp), deren Hauptfunktion die strukturelle Unterstützung der Organe ist.
Bindegewebe besteht aus drei Hauptelementen: amorphe Grundsubstanz, Fasern und Zellen. Die Art und das Verhältnis dieser Elemente bestimmen die Art des Bindegewebes.
Amorphe Grundsubstanz:
Fasern (zwei Arten):
Zellen:

Mikroskopische Darstellung eines Osteoblasten (100x; Wright-Giemsa-Färbung)
Bild: “Osteoblast (Wright-Giemsa stain, 100x)“ von Gabriel Caponetti. Lizenz: CC BY 3.0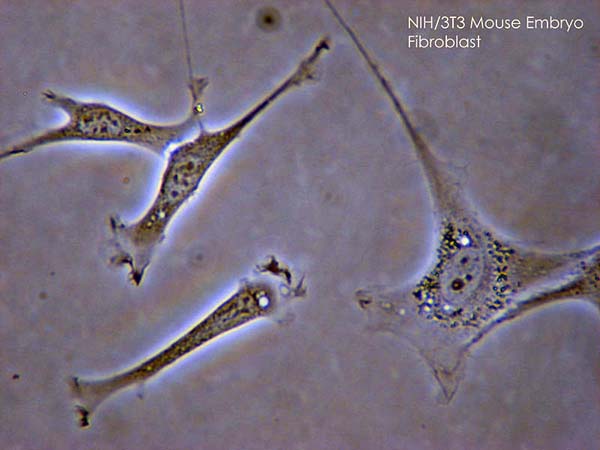
Fibroblasten (Phasenkontrastmikroskopie)
Bild: „Fibroblast“ von SubtleGuest. Lizenz: CC BY 2.5
Lockeres Bindegewebe
Bild: „Connective Tissue Loose Aerolar (41743649782)“ von Berkshire Community College Bioscience Image Library. Lizenz: CC0 1.0
Fettgewebe
Bild: „Illu connective tissues 1“ vom National Cancer Institute. Lizenz: Public Domain, bearbeitet von Lecturio.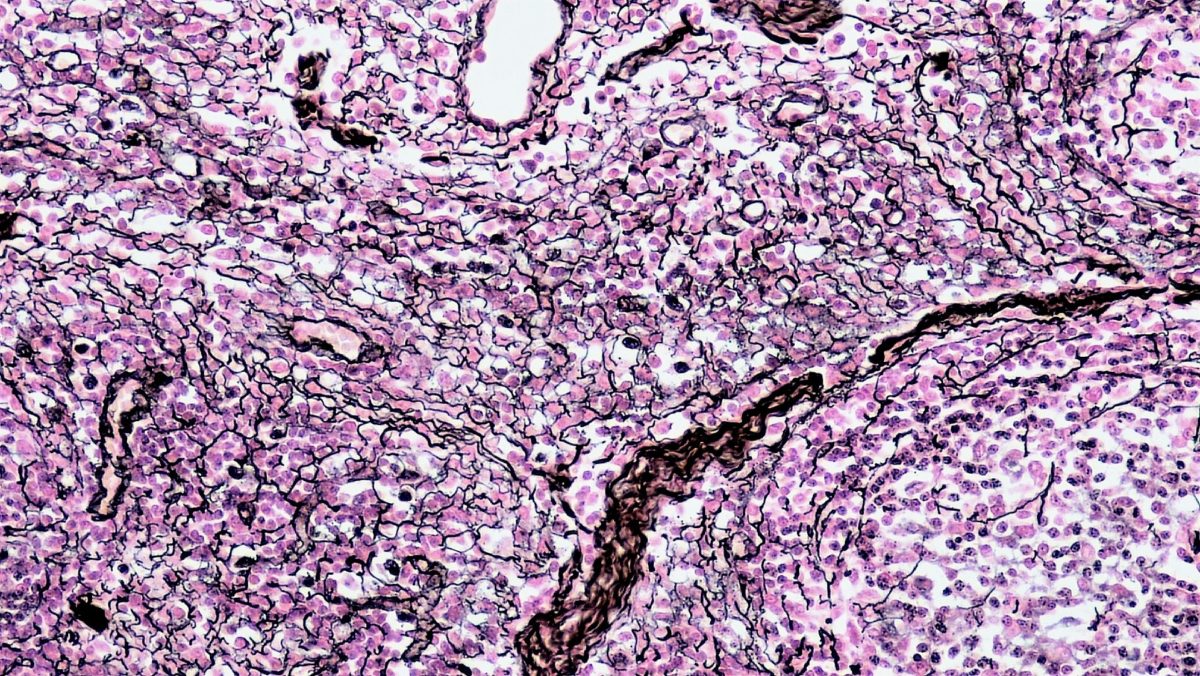
Retikuläres Bindegewebe
Bild: „Connective Tissue Reticular (40885193805)“ von Berkshire Community College Bioscience Image Library. Lizenz: CC0 1.0
Sehne (straffes parallelfaseriges Bindegewebe)
Bild: „Connective Tissue Tendon (26987380607)“ von Berkshire Community College Bioscience Image Library. Lizenz: CC0 1.0
Elastisches Bindegewebe
Bild: „Connective Tissue Human Elastic Tissue (41813163882)“ von Berkshire Community College Bioscience Image Library. Lizenz: CC0 1.0

















