Rezeptoren sind ProteineProteineProteine und Peptide auf der Oberfläche oder innerhalb einer Zelle, an die als Liganden bezeichnete Signalmoleküle (z. B. HormoneHormoneEndokrines System: Überblick) binden und eine bestimmte Reaktion innerhalb der Zelle auslösen können. Oberflächenrezeptoren sind an die ZellmembranZellmembranDie Zelle: Zellmembran gebunden, empfangen Signale aus ihrer Umgebung und übertragen diese Signale in die Zelle, oft über Second MessengerSecond MessengerSecond Messenger (z. B. cAMP) oder durch Phosphorylierungskaskaden. Die Oberflächenrezeptoren können in verschiedene Unterklassen unterteilt werden. Zu den drei wichtigsten Klassen gehören ligandengesteuerte Ionenkanalrezeptoren, Enzym-gekoppelte Rezeptoren (meist Rezeptor-Tyrosinkinasen) und G-Protein-gekoppelte Rezeptoren (GPCRs). Intrazelluläre Rezeptoren hingegen befinden sich im Zytoplasma oder Nukleus und fungieren häufig als Transkriptionsfaktoren, die direkt mit der DNADNADie Desoxyribonukleinsäure – Aufbau, Struktur und verschiedene Arten der DNA interagieren und die Genexpression beeinflussen.
Rezeptoren sind ProteineProteineProteine und Peptide auf der Oberfläche oder innerhalb einer Zelle, an die als Liganden bezeichnete Signalmoleküle (z. B. HormoneHormoneEndokrines System: Überblick) binden und eine bestimmte Reaktion innerhalb der Zelle auslösen können.
Allgemeine Physiologie
Bindung des Liganden an den Rezeptor → Konformationsänderung des Rezeptorproteins
Drei Haupttypen von Oberflächenrezeptoren (Membranrezeptoren): Ligandengesteuerte Ionenkanäle, Enzym-gekoppelte Rezeptoren und G-Protein-gekoppelte Rezeptoren (GPCRs)
Bild von Lecturio.
Intrazelluläre Rezeptoren
Innerhalb der Zelle:
Im Zytoplasma
Innerhalb des Zellkerns
Zusammengesetzt aus drei Domänen:
Ligandenbindungsdomäne
DNA-bindende Domäne
Domäne, die mit anderen Transkriptionsfaktoren interagieren kann (z. B. Coaktivatoren, Repressoren)
Liganden von intrazellulären Rezeptoren sind meist kleine hydrophobe Moleküle, die durch die ZellmembranZellmembranDie Zelle: Zellmembran diffundieren können.
Beispiele für intrazelluläre Rezeptoren:
Steroidrezeptoren
Vitamin-D-Rezeptor
Guanylatcyclase (NO-Rezeptor, der als Enzym fungiert und cGMP synthetisiert)
Intrazelluläre Rezeptoren: Intrazelluläre Rezeptoren können im Zytoplasma oder im Zellkern lokalisiert sein. Rezeptoren, die sich im Zytoplasma befinden, wandern nach Bindung ihres Liganden (z. B. Hormone) in den Zellkern. Im Zellkern bindet der Hormon-Rezeptor-Komplex an Hormone Responsive Elements (HREs), bei denen es sich um spezifische DNA-Sequenzen handelt. Der Komplex bindet weitere Transkriptionsfaktoren, um die Genexpression zu modulieren.
Ligandengesteuerte Ionenkanäle: Die Abbildung zeigt die Bindung eines Liganden an einen ligandengesteuerten Ionenkanal (grüne Struktur). Die Bindung induziert eine Konformationsänderung im Kanal, sodass Ionen hindurchtreten können.
Bild von Lecturio.
Physiologie
Bindung des Liganden (z. B. Acetylcholin) an den Ionenkanal
Konformationsänderung → Öffnung des Kanals
Ionen diffundieren entlang ihres elektrochemischen Gradienten durch den Kanal (z. B. Na+, K+, Ca2+ und/oder Cl–).
Funktionen:
Übertragung von Nervensignalen
Muskelkontraktion
Hormonausschüttung
T-Zellen-Aktivierung
Dissoziation des Liganden → Schließung des Kanals
Klinische Relevanz
Myasthenia gravisMyasthenia gravisMyasthenia gravis: eine neuromuskuläre Autoimmunerkrankung, die durch Muskelschwäche und -ermüdbarkeit gekennzeichnet ist. Ursache ist eine Dysfunktion oder Zerstörung von Acetylcholin-Rezeptoren an der motorischen Endplatte (ligandengesteuerte Ionenkanäle). Nach Bindung von Acetylcholin öffnen sich die Kanäle und ermöglichen einen Einstrom von Na+ in die Zelle, was zu einer Depolarisation führt. Folge ist eine Muskelkontraktion. Ohne funktionale Acetylcholin-Rezeptoren ist die Muskelkontraktion gestört. Myasthenia gravisMyasthenia gravisMyasthenia gravis manifestiert sich mit Müdigkeit, Ptosis, DysphagieDysphagieDysphagie, Atembeschwerden und fortschreitender Schwäche der Muskulatur in den Extremitäten, die zu Bewegungsschwierigkeiten führt.
Enzym-gekoppelte Rezeptoren
Überblick
Rezeptoren mit enzymatischer Aktivität nach Aktivierung durch ihren Liganden
Häufig Kinasen: EnzymeEnzymeGrundlagen der Enzyme, die die Übertragung einer Phosphatgruppe von einem Molekül auf ein anderes katalysieren
Transmembranhelix, die extrazelluläre und intrazelluläre Komponenten verbindet
Struktur der Rezeptor-Tyrosinkinasen (RTKs) mit drei Domänen. Liganden binden an die extrazelluläre Ligandenbindungsdomäne.
Bild von Lecturio.
Signalweg
Bindung des Liganden an die einzelnen RTK-Rezeptoren:
Dimerisierung zweier benachbarter RTKs
Autophosphorylierung:
Gegenseitige Phoyphorylierung der Dimere
Ein RTK-Monomer überträgt eine Phosphatgruppe von einem ATP auf das andere RTK-Monomer (Tyrosin wird zu Phosphotyrosin).
Phosphotyrosine fungieren als Bindungsstelle für andere ProteineProteineProteine und Peptide, die an der Signaltransduktionskaskade beteiligt sind.
Die zelluläre Antwort der RTK-Aktivierung ist abhängig von den nachgeschalteten Proteinen, die in der Signalkaskade folgen.
Vorteil einer Kinasekaskade ist die Möglichkeit der Verstärkung von Signalwirkungen innerhalb der Zelle.
Fehlfunktionen dieser Kaskade stehen mit Krebs in Verbindung, da die Regulation allgemeiner Zellfunktionen gestört wird.
Rezeptor-Tyrosinkinase-Funktion: Bindet ein Ligand an die extrazelluläre Domäne der RTK, dimerisieren zwei RTKs. Nach der Dimerisierung überträgt jede RTK eine Phosphatgruppe von ATP auf die andere RTK. Dieser Prozess wird als Autophosphorylierung bezeichnet. Nach der Autophosphorylierung fungieren die Phosphotyrosine als Bindungs- und Aktivierungsstellen für weitere Proteine, in der Regel für Enzyme. Die Art der zellulären Antwort hängt davon ab, welche Proteine in der Signalkaskade folgen. Tyr: Tyrosin; P: Phosphat.
Bild von Lecturio.
Beispiele für RTKs
Insulinrezeptoren:
Reife Insulinrezeptoren (RTKs) liegen als inaktive Dimere vor, die aus zwei α- und zwei β-Untereinheiten bestehen. Die Untereinheiten sind über Disulfidbrücken miteinander verbunden:
α-Untereinheiten: extrazelluläre Domänen mit Ligandenbindungsstelle
β-Untereinheiten: Transmembrandomänen und intrazelluläre Tyrosinkinase-Domänen
Bindung von InsulinInsulinInsulin an die α-Domänen → Autophosphorylierung
Signalweg:
Insulin-Rezeptor-Substrat 1 (IRS-1) bindet an die Phosphotyrosine →
Aktivierung der Phosphatidylinositol-3-Kinase (PI3K) →
Phosphorylierung von PIP2 zu PIP3 →
PIP3 aktiviert die PDK1 (Phosphoinositid-abhängige Kinase 1), die wiederum die Proteinkinase B (AKT) aktiviert →
Translokation von GLUT4 (Proteinkanal für die Glukoseaufnahme in die Zelle) zur Plasmamembran →
↑ Glukose kann in die Zelle eindringen
Ein weiterer Insulin-Signalweg führt zu einer erhöhten Transkription und Translation der Glykogensynthase → ↑ Umwandlung von Glukose in Glykogen innerhalb der Zelle
Der Insulinrezeptor, ein RTK: Die Bindung von Insulin löst über das Insulin-Rezeptor-Substrat (IRS) eine Phosphorylierungskaskade aus, die schließlich zur Aktivierung der Glykogensynthase führt. Diese wandelt die zusätzliche Glukose in Glykogen um.
Bild von Lecturio.
Epidermale Wachstumsfaktorrezeptoren (Englisches Akronym: EGFRs) und der RAS-Signalweg:
Bindung des epidermalen Wachstumsfaktors (Englisches Akronym: EGF) an EGFR → Dimerisierung und Autophosphorylierung
Zusammenbau eines Signalkomplexes aus Proteinen an den Phosphotyrosinen
Übertragung eine Phosphatgruppe von GTP auf RAS-GDP → Bildung von RAS-GTP (aktive Form von RAS)
RAS-Signalweg:
RAS-GTP → Aktivierung der RAF-Kinase → Aktivierung der MEK-Kinase → Aktivierung der MAP-Kinase (MAPK)
MAPK aktiviert MNK und RSK, die spezifische, für die Zellteilung erforderliche, Transkriptionsfaktoren phosphorylieren.
RAS hat eine GTPase-Aktivität:
Abspaltung einer Phosphatgruppe → RAS-GTP inaktiviert sich selbst wieder zu RAS-GDP
Die Bindung des epidermalen Wachstumsfaktors (EGF) an den EGFR induziert die Phosphorylierung von RAS.
Bild von Lecturio.
Phosphoryliertes RAS aktiviert eine Signalkaskade, die die Zellteilung stimuliert.
Bild von Lecturio.
Klinische Relevanz
Anomalien in RTKs sind Ursache einer Reihe verschiedener angeborener Fehlbildungssyndrome und Krebsarten. Dabei handelt es sich insbesondere um Gain-of-Function-Mutationen, die zu einer übermäßigen Zellproliferation führen.
Chronische myeloische Leukämie (CML): eine maligne Proliferation der granulozytären Zelllinie aufgrund einer reziproken Translokation (9;22)(q34;q11). Das ChromosomChromosomGrundbegriffe der Genetik enthält ein BCR-ABL1-Fusionsgen (aus ABL1 auf ChromosomChromosomGrundbegriffe der Genetik 9 und BCR auf ChromosomChromosomGrundbegriffe der Genetik 22), das konstitutiv aktive Tyrosinkinasen produziert. Daraus folgt eine unkontrollierte, granulozytäre Zellproliferation. Die Erkrankung kann asymptomatisch sein. Betroffene können aber auch unter unspezifischen Allgemeinsymptomen, Schmerzen im Bereich des Sternums und einer SplenomegalieSplenomegalieSplenomegalie leiden.
Achondroplasie: eine autosomal-dominant vererbte Knochendysplasie aufgrund von Gain-of-Function-Mutationen im Fibroblasten-Wachstumsfaktor-Rezeptor-3-Gen (FGFR3-Gen), das für eine RTK kodiert. Als Folge wird der FGFR3-Rezeptor permanent aktiviert, was die Proliferation von ChondrozytenChondrozytenKnorpelgewebe hemmt und zu einer gestörten Knochenbildung und Skelettanomalien führt.
G-Protein-gekoppelte Rezeptoren
Überblick
G-Protein-gekoppelte Rezeptoren (Englisches Akronym: GPCRs) sind Transmembranproteine, die auf ihrer extrazellulären Seite Hormonliganden binden. Die Ligandenbindung induziert eine Konformationsänderung der intrazellulären Domäne, sodass ein assoziiertes G-Protein aktiviert wird, das dann über Second MessengerSecond MessengerSecond Messenger eine Signalkaskade auslöst.
Größte Rezeptorfamilie mit über 800 identifizierten GPCR-Genen
An vielen metabolischen und strukturellen Veränderungen innerhalb der Zelle beteiligt
Beispiele:
β-adrenerge Rezeptoren
Geruchsrezeptoren
Struktur
Extrazelluläre Domäne mit Ligandenbindungsstelle
Transmembrandomäne:
Verankerung des Rezeptors in der Plasmamembran
Häufig sieben α-Helices (7-Transmembranhelix-Rezeptoren (7-TM-Rezeptoren))
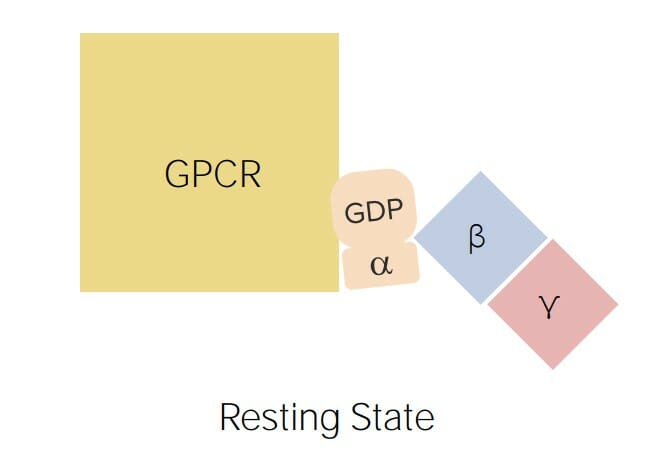
Bindung der Guaninnukleotide: GTP (aktiver Zustand) und GDP (inaktiver Zustand)
Hydrolyse von GTP zu GDP → nach Aktivierung durch GTP inaktiviert sich die α-Untereinheit schließlich selbst durch hydrolytische Umwandlung von GTP in GDP
β und γ: Unterstützung der Bindung des α‒GDP mit dem GPCR
G-Protein-gekoppelter Rezeptor (GPCR): Die G-Proteine bestehen aus drei Untereinheiten: α (in seiner inaktiven Form an GDP und in seiner aktiven Form an GTP gebunden), β und γ (unterstützen die Assoziation mit dem GPCR).
Bild von Lecturio.
Aktivierungsweg
Bindung des Liganden an die Bindungsstelle der extrazellulären Domäne des Rezeptors → Konformationsänderung im GPCR
Phosphorylierung des an die α-Untereinheit gebundenen GDP → α‒GTP (aktiviertes G-Protein)
α‒GTP trennt sich von der β- und γ-Untereinheit → Phosphorylierung des nächsten Proteins der Signalkaskade (Effektorprotein)
Die Phosphorylierung kann aktivierend oder inaktivierend wirken.
Synthese von Second Messengern
Aktivierungsweg G-Protein-gekoppelter Rezeptoren:
Ein Ligand bindet an den GPCR und induziert intern eine Konformationsänderung. Diese Konformationsänderung bewirkt, dass die α-Untereinheit des G-Proteins ein GDP gegen ein GTP austauscht. Dadurch wird das G-Protein aktiviert. Die GTP-gebundene α-Untereinheit trennt sich von den β- und γ-Untereinheiten und aktiviert ein Effektorenzym (über Phosphorylierung vom GTP). Das Effektorenzym aktiviert anschließend einen Second Messenger (hier cAMP), der das Signal innerhalb der Zelle weiterleitet.
Bild von Lecturio.
Adenylatzyklase (AC) und das cAMP-Second-Messenger-System
Die Adenylatcyclase bzw. Adenylylcyclase (AC) ist ein verbreitetes GPCR-Effektorprotein. Das aktivierte G-Protein phosphoryliert und aktiviert die AC. Die AC wandelt ATP in cAMP um. cAMP ist ein wichtiger Second MessengerSecond MessengerSecond Messenger.
Beispiel 1: Glucagon-Signalweg
Bindung von Glucagon an den GPCR → Konformationsänderung
Die zytosolisch gebundene α-Untereinheit des G-Proteins tauscht GDP gegen GTP aus und trennt sich von den β- und γ-Untereinheiten.
α‒GTP (aktive Form) phosphoryliert und aktiviert die AC.
AC synthetisiert cAMP aus ATP.
cAMP phosphoryliert die regulatorischen Untereinheiten der Proteinkinase A (PKA) → Ablösung der regulatorischen Untereinheiten von den katalytischen Untereinheiten (Aktivierung der PKA)
Wirkung der PKA:
Inaktivierung der Glykogensynthase → Hemmung der Glykogensynthese
Aktivierung der Phosphorylase-Kinase (PK) → Aktivierung der Glykogen-Phosphorylase durch die PK → Stimulation des Glykogenabbaus
Zusammenfassung: Glucagon stimuliert den Glykogenabbau zu Glukose und hemmt gleichzeitig die Glykogensynthese.
G-Protein-gekoppelter Rezeptor (GPCR) gekoppelt an die Adenylatcyclase (AC):
Aktive Enzyme sind in Grün dargestellt, inaktive Enzyme sind in Rot abgebildet. AC wandelt ATP in cAMP um, das wiederum die Proteinkinase A (PKA) aktiviert. PKA phosphoryliert und inaktiviert sowohl die Glykogen-Synthase (GS), als auch die Phosphorylase-Kinase (PK), die dadurch aktiviert wird. Die PK aktiviert die Glykogenphosphorylase, die den Glykogenabbau zu Glukose stimuliert.
GTP: Guanosintriphosphat
GDP: Guanosindiphosphat
ATP: Adenosintriphosphat
cAMP: zyklisches Adenosinmonophosphat
Bild von Lecturio
Beispiel 2: Adrenalin-Signalwege (unterschiedliche Wirkung je nach in der Zielzelle vorhandener ProteineProteineProteine und Peptide)
Darm: Bindung von Adrenalin an einen GPCR → Hemmung der AC → Entspannung der Darmmuskulatur und Verlangsamung der VerdauungVerdauungDigestion und Resorption
Zwei verschiedene G-Proteine können denselben intrazellulären Signalübertragungsweg aktivieren: Sowohl Adrenalin als auch Glucagon stimulieren den Glykogenabbau in Glukose.
Bild von Lecturio.
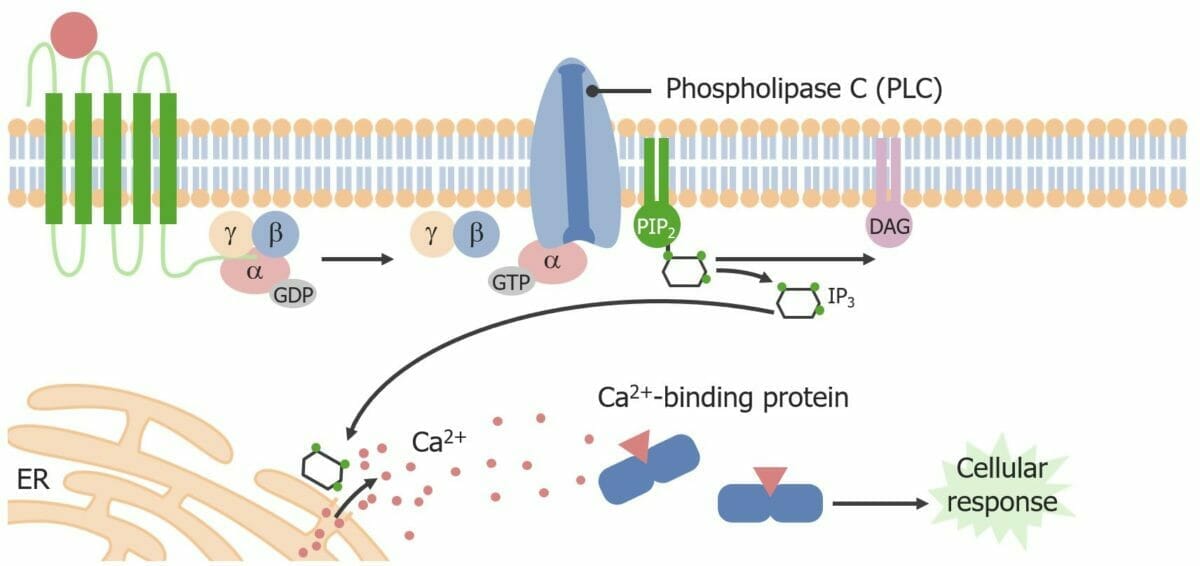
Phospholipase C (Effektorprotein) und die sekundären Botenstoffe IP3 und DAG
Ligandenbindung an den GPCR → Konformationsänderung
Die α-Untereinheit des G-Proteins tauscht GDP gegen GTP aus und trennt sich von den β- und γ-Untereinheiten.
α‒GTP (aktiv) phosphoryliert und aktiviert die Phospholipase C (PLC).
Ca2+-abhängige Aktivierung der Proteinkinase C (PKC) durch DAG → Phosphorylierung weiterer ProteineProteineProteine und Peptide
Beispiel:
Bindung von Acetylcholin (ACh) an einen muskarinischen GPCR in Belegzellen des Magens → Aktivierung der PLC → Synthese von IP3 und DAG → Ca2+ aus dem ER und DAG aktivieren die H+/K+-ATPase, die H + Ionen in das Magenlumen transportiert
GPCR aktiviert Phospholipase C, die PIP2 (Phosphatidylinositol-4,5-Bisphosphat) in IP3 und DAG umwandelt. IP3 bindet an einen ligandengesteuerten Ionenkanal am endoplasmatischen Retikulum. Durch Öffnung des Kanals wird Calcium in das Zytoplasma freigesetzt. Das induziert eine zelluläre Reaktion (z. B. Hormonfreisetzung aus endokrinen Zellen).
Bild von Lecturio.
Klinische Relevanz
Über 30 verschiedene menschliche Erkrankungen sind auf Mutationen in GPCRs zurückzuführen. Diese Mutationen können aktivierend oder inaktivierend wirken. Ein Beispiel ist der DiabetesDiabetesDiabetes Mellitus insipidus renalis.
DiabetesDiabetesDiabetes Mellitus insipidus renalis: eine durch Mutationen des V2-Rezeptors verursachte Erkrankung, die zu einer ADH-Resistenz (antidiuretisches Hormon) führt. Der V2-Rezeptor ist ein GPCR, über den normalerweise der Einbau von Aquaporinen in die Membranen von renalen Sammelrohrzellen vermittelt und dadurch die Wasserresorption ermöglicht wird. Aufgrund der Mutation des GPCR wird das ADH-Signal ineffektiv innerhalb der Zelle übertragen. Die unzureichende Wirkung von ADH resultiert in einer Asthenurie.
Nukleäre Rezeptoren
Überblick
Subtyp der intrazellulären Rezeptors
Wirkung als Liganden-abhängige Transkriptionsfaktoren, die die Genexpression modulieren
Obwohl sie als nukleäre Rezeptoren bezeichnet werden, sind sie häufig im Zytoplasma lokalisiert und wandern nach Ligandenbindung in den Zellkern.
Interaktion mit anderen Transkriptionsfaktoren → Modulation der Genexpression (Wirkung als Enhancer oder Repressor)
Aktivierungsweg nukleärer Rezeptoren
Bild von Kevin Ahern, MD.
Klinische Relevanz
Androgeninsensitivitätssyndrom (AIS): eine X-chromosomal rezessive Erkrankung, bei der eine genetische Mutation die Funktion von Androgenrezeptoren beeinflusst und zu Testosteronresistenz führt. Die Androgenrezeptoren sind nukleäre Rezeptoren, die im Zytoplasma lokalisiert sind, nach Bindung an AndrogeneAndrogeneAndrogene und Antiandrogene in den Zellkern wandern und die Transkription von Synthese von Proteinen mit androgenen Wirkungen erhöhen. Betroffene Personen haben einen 46,XY-Karyotyp und Hodenhochstand, bei gleichzeitiger Ausprägung weiblicher Genitalien und Brustentwicklung (aufgrund der peripheren Umwandlung des überschüssigen Testosterons in Östrogen).
O’Shea, JJ, et al. (2015). The Jak-STAT pathway: Impact on human disease and therapeutic intervention. Annual Review of Medicine. Abgerufen am 26. Oktober 2021 von https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5634336/
McDonell, LM, Kernohan, KD, Boycott, KM und Sawyer, SL (2015). Receptor tyrosine kinase mutations in developmental syndromes and cancer: Two sides of the same coin. Human Molecular Genetics, 24(R1), R60–R66. https://doi.org/10.1093/hmg/ddv254
Lecturio und die TÜV SÜD Akademie erhielten für den gemeinsam entwickelten Online-Kurs zur Vorbereitung auf den Drohnenführerschein den eLearning Award 2019 in der Kategorie “Videotraining”.
Comenius-Award 2019
Die Lecturio Business Flat erhielt 2019 das Comenius-EduMedia-Siegel, mit dem die Gesellschaft für Pädagogik, Information und Medien jährlich pädagogisch, inhaltlich und gestalterisch herausragende didaktische Multimediaprodukte auszeichnet.
IELA-Award 2022
Die International E-Learning Association, eine Gesellschaft für E-Learning Professionals und Begeisterte, verlieh der Lecturio Learning Cloud die Gold-Auszeichnung in der Kategorie “Learning Delivery Platform”.
Comenius-Award 2022
In der Kategorie “Lehr- und Lernmanagementsysteme” erhielt die Lecturio Learning Cloud die Comenius-EduMedia-Medaille. Verliehen wird der Preis von der Gesellschaft für Pädagogik, Information und Medien für pädagogisch, inhaltlich und gestalterisch herausragende Bildungsmedien.
B2B Award 2020/2021
Die Deutsche Gesellschaft für Verbraucherstudien (DtGV) hat Lecturio zum Branchen-Champion unter den deutschen Online-Kurs-Plattformen gekürt. Beim Kundenservice belegt Lecturio den 1. Platz, bei der Kundenzufriedenheit den 2. Platz.
B2B Award 2022
Für herausragende Kundenzufriedenheit wurde Lecturio von der Deutschen Gesellschaft für Verbraucherstudien (DtGV) mit dem deutschen B2B-Award 2022 ausgezeichnet. In der Rubrik Kundenservice deutscher Online-Kurs-Plattformen belegt Lecturio zum zweiten Mal in Folge den 1. Platz.
Simon Veiser
Simon Veiser beschäftigt sich seit 2010 nicht nur theoretisch mit IT Service Management und ITIL, sondern auch als leidenschaftlicher Berater und Trainer. In unterschiedlichsten Projekten definierte, implementierte und optimierte er erfolgreiche IT Service Management Systeme. Dabei unterstützte er das organisatorische Change Management als zentralen Erfolgsfaktor in IT-Projekten. Simon Veiser ist ausgebildeter Trainer (CompTIA CTT+) und absolvierte die Zertifizierungen zum ITIL v3 Expert und ITIL 4 Managing Professional.
Dr. Frank Stummer
Dr. Frank Stummer ist Gründer und CEO der Digital Forensics GmbH und seit vielen Jahren insbesondere im Bereich der forensischen Netzwerkverkehrsanalyse tätig. Er ist Mitgründer mehrerer Unternehmen im Hochtechnologiebereich, u.a. der ipoque GmbH und der Adyton Systems AG, die beide von einem Konzern akquiriert wurden, sowie der Rhebo GmbH, einem Unternehmen für IT-Sicherheit und Netzwerküberwachung im Bereich Industrie 4.0 und IoT. Zuvor arbeitete er als Unternehmensberater für internationale Großkonzerne. Frank Stummer studierte Betriebswirtschaft an der TU Bergakademie Freiberg und promovierte am Fraunhofer Institut für System- und Innovationsforschung in Karlsruhe.
Sobair Barak
Sobair Barak hat einen Masterabschluss in Wirtschaftsingenieurwesen absolviert und hat sich anschließend an der Harvard Business School weitergebildet. Heute ist er in einer Management-Position tätig und hat bereits diverse berufliche Auszeichnungen erhalten. Es ist seine persönliche Mission, in seinen Kursen besonders praxisrelevantes Wissen zu vermitteln, welches im täglichen Arbeits- und Geschäftsalltag von Nutzen ist.
Wolfgang A. Erharter
Wolfgang A. Erharter ist Managementtrainer, Organisationsberater, Musiker und Buchautor. Er begleitet seit über 15 Jahren Unternehmen, Führungskräfte und Start-ups. Daneben hält er Vorträge auf Kongressen und Vorlesungen in MBA-Programmen. 2012 ist sein Buch „Kreativität gibt es nicht“ erschienen, in dem er mit gängigen Mythen aufräumt und seine „Logik des Schaffens“ darlegt. Seine Vorträge gestaltet er musikalisch mit seiner Geige.
Holger Wöltje
Holger Wöltje ist Diplom-Ingenieur (BA) für Informationstechnik und mehrfacher Bestseller-Autor. Seit 1996 hat er über 15.800 Anwendern in Seminaren und Work-shops geholfen, die moderne Technik produktiver einzusetzen. Seit 2001 ist Holger Wöltje selbstständiger Berater und Vortragsredner. Er unterstützt die Mitarbeiter von mittelständischen Firmen und Fortune-Global-500- sowie DAX-30-Unternehmen dabei, ihren Arbeitsstil zu optimieren und zeigt Outlook-, OneNote- und SharePoint-Nutzern, wie sie ihre Termine, Aufgaben und E-Mails in den Griff bekommen, alle wichtigen Infos immer elektronisch parat haben, im Team effektiv zusammenarbeiten, mit moderner Technik produktiver arbeiten und mehr Zeit für das Wesentliche gewinnen.
Frank Eilers
Frank Eilers ist Keynote Speaker zu den Zukunftsthemen Digitale Transformation, Künstliche Intelligenz und die Zukunft der Arbeit. Er betreibt seit mehreren Jahren den Podcast „Arbeitsphilosophen“ und übersetzt komplexe Zukunftsthemen für ein breites Publikum. Als ehemaliger Stand-up Comedian bringt Eilers eine ordentliche Portion Humor und Lockerheit mit. 2017 wurde er für seine Arbeit mit dem Coaching Award ausgezeichnet.
Yasmin Kardi
Yasmin Kardi ist zertifizierter Scrum Master, Product Owner und Agile Coach und berät neben ihrer Rolle als Product Owner Teams und das höhere Management zu den Themen agile Methoden, Design Thinking, OKR, Scrum, hybrides Projektmanagement und Change Management.. Zu ihrer Kernkompetenz gehört es u.a. internationale Projekte auszusteuern, die sich vor allem auf Produkt-, Business Model Innovation und dem Aufbau von Sales-Strategien fokussieren.
Leon Chaudhari
Leon Chaudhari ist ein gefragter Marketingexperte, Inhaber mehrerer Unternehmen im Kreativ- und E-Learning-Bereich und Trainer für Marketingagenturen, KMUs und Personal Brands. Er unterstützt seine Kunden vor allem in den Bereichen digitales Marketing, Unternehmensgründung, Kundenakquise, Automatisierung und Chat Bot Programmierung. Seit nun bereits sechs Jahren unterrichtet er online und gründete im Jahr 2017 die „MyTeachingHero“ Akademie.
Andreas Ellenberger
Als akkreditierter Trainer für PRINCE2® und weitere international anerkannte Methoden im Projekt- und Portfoliomanagement gibt Andreas Ellenberger seit Jahren sein Methodenwissen mit viel Bezug zur praktischen Umsetzung weiter. In seinen Präsenztrainings geht er konkret auf die Situation der Teilnehmer ein und erarbeitet gemeinsam Lösungsansätze für die eigene Praxis auf Basis der Theorie, um Nachhaltigkeit zu erreichen. Da ihm dies am Herzen liegt, steht er für Telefoncoachings und Prüfungen einzelner Unterlagen bzgl. der Anwendung gern zur Verfügung.
Zach Davis
Zach Davis ist studierter Betriebswirt und Experte für Zeitintelligenz und Zukunftsfähigkeit. Als Unternehmens-Coach hat er einen tiefen Einblick in über 80 verschiedene Branchen erhalten. Er wurde 2011 als Vortragsredner des Jahres ausgezeichnet und ist bis heute als Speaker gefragt. Außerdem ist Zach Davis Autor von acht Büchern und Gründer des Trainingsinstituts Peoplebuilding.
Wladislav Jachtchenko
Wladislaw Jachtchenko ist mehrfach ausgezeichneter Experte, TOP-Speaker in Europa und gefragter Business Coach. Er hält Vorträge, trainiert und coacht seit 2007 Politiker, Führungskräfte und Mitarbeiter namhafter Unternehmen wie Allianz, BMW, Pro7, Westwing, 3M und viele andere – sowohl offline in Präsenztrainings als auch online in seiner Argumentorik Online-Akademie mit bereits über 52.000 Teilnehmern. Er vermittelt seinen Kunden nicht nur Tools professioneller Rhetorik, sondern auch effektive Überzeugungstechniken, Methoden für erfolgreiches Verhandeln, professionelles Konfliktmanagement und Techniken für effektives Leadership.
Alexander Plath
Alexander Plath ist seit über 30 Jahren im Verkauf und Vertrieb aktiv und hat in dieser Zeit alle Stationen vom Verkäufer bis zum Direktor Vertrieb Ausland und Mediensprecher eines multinationalen Unternehmens durchlaufen. Seit mehr als 20 Jahren coacht er Führungskräfte und Verkäufer*innen und ist ein gefragter Trainer und Referent im In- und Ausland, der vor allem mit hoher Praxisnähe, Humor und Begeisterung überzeugt.