Die DNA ist das Molekül, in der die gesamte genetische Information des Menschen gespeichert bzw. kodiert ist. Beim Menschen ist die DNA in Form von 23 Chromosomenpaaren im Zellkern organisiert. Sie stellt die grundlegende Matrize (Vorlage) für die Replikation genetischer Informationen, die RNA-Transkription und die Proteinbiosynthese dar. Die DNA ist folglich die Basis all unserer Zellfunktionen und somit essentiell für das Überleben.
Kostenloser
Download
Lernleitfaden
Medizin ➜
Die DNA ist eine doppelsträngige Helix. Die DNA-Bestandteile sind die Nukleotide:

Struktur der Desoxyribonukleotide
Bild von Lecturio.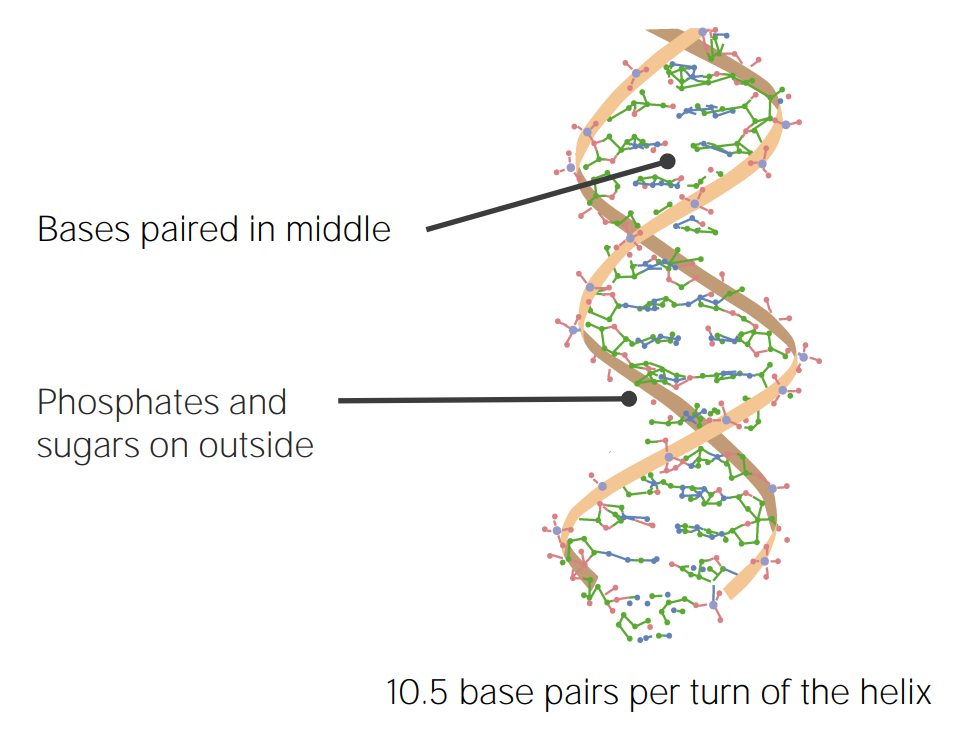
DNA-Struktur nach Watson und Crick
Bild von Lecturio.
Aufbau der DNA-Doppelhelix, der die antiparallele (polare) Orientierung jedes komplementären Strangs zeigt
Bild von Lecturio.
DNA-Struktur und die 2 Formen des Chromatins:
Euchromatin (aktiv), wobei DNA repliziert oder transkribiert wird, und Heterochromatin (inaktiv), wobei DNA nicht repliziert oder transkribiert wird.


















